



Sommario
- Introduzione
- Stadi 1-4. Fase giovanile: sviluppo in altezza e costruzione del tronco
- Stadi 5 e 6. Fase adulta: espansione in larghezza
- Stadi 7 e 8. Fase di maturità: strategia di durata
- Stadi 9 e 10. Fase di senescenza: riduzione della chioma
- Lo sviluppo degli alberi: logica e implicazioni
Introduzione
Ogni albero ha un proprio portamento, una fisionomia, un’architettura (disposizione dei rami nello spazio e loro sviluppo relativo) che lo rendono spesso riconoscibile anche a distanza.
Sebbene questa architettura, risultato di una crescita del tronco e dei rami non uniforme nel tempo, sia geneticamente programmata per ciascuna specie, Raimbault ha descritto uno schema generale valido per tutti gli alberi, suddivisibile in 10 stadi di sviluppo morfofisiologico (Fig. 1).
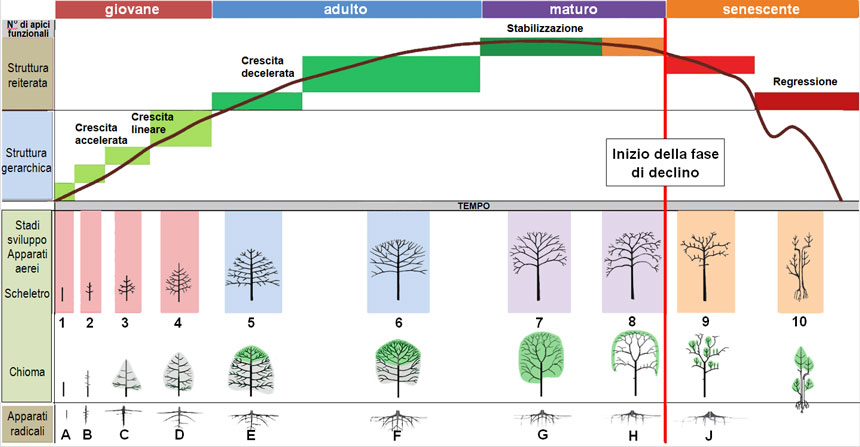 Fig. 1. Schema di sviluppo naturale della struttura arborea, in base all’evoluzione morfologica delle sue componenti aerea (stadi 1-10) e sotterranea (stadi A-J). La curva mostra l’andamento del numero di apici (nelle ordinate) nel corso del tempo (nelle ascisse): per le successive ramificazioni, il numero aumenta fino alla fase adulta, si stabilizza alla maturità per scendere poi rapidamente nella senescenza. Gli apparati aerei sono rappresentati in due righe: la prima mostra il tronco e le ramificazioni, la seconda la chioma (in grigio quella temporanea, in verde quella definitiva); si noti che allo stadio 8 la chioma forma solo uno strato periferico, mentre l’interno è privo di foglie. Nella fase giovanile (stadi 1-4) l’albero si sviluppa in altezza, assumendo una forma piramidale. Nella fase adulta (stadi 5-6) raggiunge le dimensioni massime, espandendosi in larghezza e arrotondando la chioma. La maturità (stadi 7-8) è una fase di mantenimento della chioma. Nella senescenza (stadi 9-10) la chioma si riduce e il tronco diviene cavo; spesso alcune branche si rendono indipendenti, con un proprio apparato radicale.
Fig. 1. Schema di sviluppo naturale della struttura arborea, in base all’evoluzione morfologica delle sue componenti aerea (stadi 1-10) e sotterranea (stadi A-J). La curva mostra l’andamento del numero di apici (nelle ordinate) nel corso del tempo (nelle ascisse): per le successive ramificazioni, il numero aumenta fino alla fase adulta, si stabilizza alla maturità per scendere poi rapidamente nella senescenza. Gli apparati aerei sono rappresentati in due righe: la prima mostra il tronco e le ramificazioni, la seconda la chioma (in grigio quella temporanea, in verde quella definitiva); si noti che allo stadio 8 la chioma forma solo uno strato periferico, mentre l’interno è privo di foglie. Nella fase giovanile (stadi 1-4) l’albero si sviluppa in altezza, assumendo una forma piramidale. Nella fase adulta (stadi 5-6) raggiunge le dimensioni massime, espandendosi in larghezza e arrotondando la chioma. La maturità (stadi 7-8) è una fase di mantenimento della chioma. Nella senescenza (stadi 9-10) la chioma si riduce e il tronco diviene cavo; spesso alcune branche si rendono indipendenti, con un proprio apparato radicale.Figura ottenuta combinando illustrazioni di Raimbault e Tanguy 1993, Sgherzi et al., 2016 e Morelli e Raimbault, 2017.
Morelli G., Raimbault P., 2017. Legami inscindibili. Acer, 2/2017: 25-32.
Raimbault P., Tanguy M., 1993. La gestion des arbres d’ornement, Rev. For. Fr. XLV (2).
Sgherzi R., Bistoni A., Puddu G., 2016. Gazzetta Ambiente, 3/2016: 109-128.
Per avere una visione d’insieme, i 10 stadi di sviluppo possono essere raggruppati in 4 fasi, ciascuna caratterizzata da una diversa strategia.
- Fase giovanile (stadi 1-4): è il periodo di formazione, caratterizzato dalla strategia di sviluppo in altezza e di costruzione del tronco. Questi stadi sono riconoscibili per la presenza di un ramo terminale del tronco (detto “freccia”) caratterizzato da una notevole crescita, che domina lo sviluppo dei rami inferiori.
- Fase adulta (stadi 5 e 6): strategia di costruzione della chioma definitiva, grazie all’espansione in larghezza, e di acquisizione della dimensione massima.
- Fase di maturità (stadi 7 e 8): strategia di durata, caratterizzata dal mantenimento del volume complessivo della chioma, grazie al rinnovamento delle unità architettoniche che la compongono.
- Fase di senescenza (stadi 9 e 10): riduzione della chioma (per morte dei rami apicali) e tentativi di ricostruzione di più tronchi funzionalmente indipendenti, sebbene appartenenti allo stesso tronco.
Stadi 1-4: sviluppo in altezza e costruzione del tronco (Fig. 2)
Stadio 1: la strategia della giovane plantula nata dal seme è la crescita in altezza del fusto; la vigorosa crescita dell’asse verticale alla ricerca della luce, infatti, è di primaria importanza per innalzarsi sullo strato erbaceo (e, nel bosco, per superare la concorrenza di altri alberi).
La crescita si realizza producendo in modo iterativo, uno sull’altro, moduli architettonici e funzionali elementari (fitomeri) costituiti da un nodo (punto di inserimento della foglia), una gemma ascellare (che in seguito svilupperà un ramo laterale) e un internodo (tratto di fusto compreso tra due nodi).
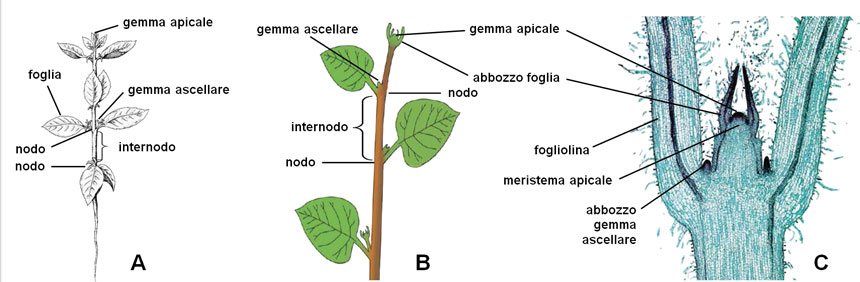 Fig. 2. A: stadio 1: la giovane plantula cresce in altezza, senza ramificarsi, per successiva formazione di nodi (nei quali sono inserite le foglie e, alla loro ascella, gemme che l’anno successivo daranno origine a rami laterali) e internodi (tratti di fusto compresi tra due nodi). B: schema del tratto apicale del fusto. C: microfotografia di una sezione longitudinale della gemma apicale; sono visibili la gemma e il suo meristema, gli abbozzi fogliari che la proteggono, due giovani foglie (la linea scura sono fasci conduttori) e due abbozzi di gemme ascellari. L’intero accrescimento in altezza è opera della gemma apicale.
Fig. 2. A: stadio 1: la giovane plantula cresce in altezza, senza ramificarsi, per successiva formazione di nodi (nei quali sono inserite le foglie e, alla loro ascella, gemme che l’anno successivo daranno origine a rami laterali) e internodi (tratti di fusto compresi tra due nodi). B: schema del tratto apicale del fusto. C: microfotografia di una sezione longitudinale della gemma apicale; sono visibili la gemma e il suo meristema, gli abbozzi fogliari che la proteggono, due giovani foglie (la linea scura sono fasci conduttori) e due abbozzi di gemme ascellari. L’intero accrescimento in altezza è opera della gemma apicale.(Microfotografia da Ugo Capasso, 2014. Il germoglio, la parte epigea della pianta. https://slideplayer.it/slide/189099/).
Affinché la crescita in altezza sia massima occorre che tutte le energie siano concentrate nella costruzione del fusto, anziché disperdersi nella costruzione di rami laterali. Ciò si ottiene grazie alla dominanza apicale, un meccanismo ormonale geneticamente preordinato che inibisce la crescita delle gemme sottostanti e assicura così che la massima quantità di acqua e nutrienti assorbita dalle radici sia recapitata alla gemma apicale.
L’apice del fusto, infatti, a livello delle foglioline, produce l’ormone auxina Auxina: dal greco auxánein ‘crescere’: ormone vegetale che regola l’accrescimento in vari organi vegetativi (germogli, radici, frutti). Chimicamente è l’acido indol-3-acetico (IAA. Indol Acetic Acid). che, da una parte, stimola la crescita del germoglio apicale (favorendo la divisione cellulare, la distensione e l’allungamento delle cellule dell’apice) mentre, dall’altra, trasportato attivamente verso il basso, raggiunge le gemme laterali sottostanti inibendone la crescita (mantenendo bassi i livelli di citochinine –gli ormoni promotori della crescita di queste gemme– provenienti dalle radici).
In sintesi, le cellule del tessuto meristematico Meristema: dal greco meristós ‘diviso’. Tessuto vegetale costituito da cellule indifferenziate dotate di un’elevata capacità di dividersi generando così nuove cellule che consentono l’accrescimento e, differenziandosi, formano i tessuti definitivi.
Il meristema primario o apicale è responsabile dell’accrescimento in lunghezza del fusto e delle radici.
Il meristema secondario o laterale (cambio) è responsabile dell’accrescimento secondario (in diametro) nel fusto e radici delle Dicotiledoni legnose e Gimnosperme. della gemma apicale (costituito da cellule totipotenti, cioè capaci di generare altre cellule differenziate: legnose, cribrose, fibrose, epidermiche, parenchimatiche) si moltiplicano ripetutamente producendo cellule figlie che, allungandosi, fanno crescere il germoglio apicale, mentre le gemme laterali sottostanti vengono mantenute in uno stato di dormienza o di crescita ridotta. Se, infatti, si taglia la gemma apicale, l’effetto inibitorio viene meno e le gemme sottostanti si sviluppano rapidamente.
Allo stadio aereo 1 corrisponde lo stadio radicale A. Anche l’apice radicale esercita la dominanza sugli abbozzi delle radici laterali, il cui sviluppo viene perciò inibito: si forma pertanto una radice a fittone, senza ramificazioni.
Va precisato che questo schema di sviluppo in 10 stadi vale per gli alberi. Negli arbusti e nelle piante erbacee (annuali e perenni), invece, la dominanza apicale non si verifica: pertanto, proprio per questo, le gemme laterali possono crescere fin dal primo anno e formare parecchi fusti. Se, infatti, una pianta replica semplicemente innumerevoli volte i propri organi fondamentali (foglia, fusto, radici di crescita e radici di assorbimento) si otterrà un arbusto con parecchi fusti (come il nocciolo) o uno sviluppo erbaceo, sul tipo delle graminacee, che resterà sulla superficie del terreno.
Perché la ripetizione degli organi elementari possa produrre una struttura di grandi dimensioni come un albero, bisogna che la pianta strutturi i suoi organi in forma gerarchica (altrimenti si formerebbe una struttura ramificata in maniera completamente anarchica). Il compito di stabilire una gerarchia (quali assi debbano svilupparsi e quando) è svolto principalmente dalla dominanza apicale che, come vedremo, sarà esercitata nuovamente dall’apice dei rami principali.
Negli stadi 2, 3 e 4 (Fig. 3), ciascuno dei quali può protrarsi diversi anni, la ramificazione si sviluppa sotto l’influenza della dominanza apicale. A differenza di quanto avviene nel primo anno (allo stadio 1), negli stadi successivi la dominanza non impedisce più lo sviluppo delle gemme laterali del fusto, ma ai nuovi rami è consentita una crescita moderata e rivolta verso l’esterno, non verso l’alto (si evita così la loro concorrenza con la cima del tronco, che continua a crescere più velocemente). Si formano pertanto branche orizzontali o leggermente inclinate verso l’alto: si parla di crescita plagiotropica o di rami plagiotropiPlagiotropo: dal greco plàgios ‘obliquo, trasverso’ e -tropo ‘rivolto’; quindi un ramo plagiotropo è perpendicolare al tronco o un po’ obliquo.).
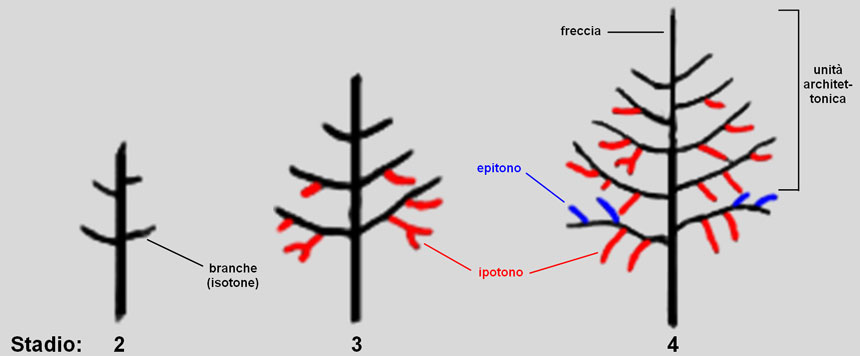 Fig. 3. Negli stadi 2, 3 e 4 la forte dominanza apicale esercitata dalla freccia modera la crescita delle branche e impedisce loro di dirigersi verso l’alto. Le branche stesse inibiscono lo sviluppo delle gemme poste sul lato superiore e ramificano pertanto sul lato inferiore (ipotonia). Verso la fine dello stadio 4, la dominanza apicale si indebolisce; ciò permette, sulle branche più basse, lo sviluppo di rami epitoni (sul lato superiore delle branche).
Fig. 3. Negli stadi 2, 3 e 4 la forte dominanza apicale esercitata dalla freccia modera la crescita delle branche e impedisce loro di dirigersi verso l’alto. Le branche stesse inibiscono lo sviluppo delle gemme poste sul lato superiore e ramificano pertanto sul lato inferiore (ipotonia). Verso la fine dello stadio 4, la dominanza apicale si indebolisce; ciò permette, sulle branche più basse, lo sviluppo di rami epitoni (sul lato superiore delle branche).
Nello stadio 2, dunque, sul vigoroso fusto verticale dominante si sviluppano ramificazioni laterali corte e suborizzontali (Fig. 3). La modalità di ramificazione è detta isotonicaRami isotoni (dal greco isos ‘uguale’): rami disposti attorno al ramo genitore, su tutti i suoi lati.
Ipotoni (dal greco hypó e dal latino scientifico hipo- ‘sotto’): rami che si sviluppano sul lato inferiore del genitore.
Epitoni (dal greco epi- ‘sopra, in, di più’): rami che si sviluppano sul lato superiore del genitore.
Anfitoni (dal greco amphí- ‘intorno, da entrambe le parti, doppio’): rami che si sviluppano (sullo stesso piano orizzontale) su entrambi i lati del genitore. poiché i rami figli sono disposti su tutti i lati del tronco.
Nello stadio 3 le branche, sempre fortemente dominate, impongono esse stesse uno sviluppo differenziato ai loro germogli ascellari, inibendo quelli sul lato superiore; si sviluppano quindi principalmente dapprima rami anfitoni (cioè posti orizzontalmente, a destra e a sinistra del ramo genitore) e poi ipotoni (cioè rivolti verso il basso).
Solo verso la fine dello stadio 4 la dominanza apicale, ancora forte sui rami alti, inizia a indebolirsi, soprattutto sulle branche più distanti dall’apice del fusto sulle quali, pertanto, possono svilupparsi anche rami epitoni (cioè sul loro lato superiore). A questo stadio risulta completata l’unità architetturale (o architettonica), rappresentata dall’estremità superiore del tronco principale (Fig. 3, stadio 4); le branche più basse, rimaste soggette più a lungo all’influenza inibitoria della dominanza apicale, andranno incontro al disseccamento e saranno eliminate per auto-potatura, denudando così la parte inferiore del tronco.
In sintesi, negli stadi di sviluppo da 1 a 4 la crescita degli alberi è fortemente influenzata dalla dominanza apicale. Per questo motivo un albero giovane è caratterizzato da un unico asse verticale (la “freccia” apicale) con una crescita molto accentuata e dai rami sottostanti, fortemente dominati da questo. Le branche laterali più vicine all’apice sono un po’ inclinate verso l’alto mentre, via via che si scende, troviamo branche che –subendo la dominanza da molti anni– saranno sempre più orizzontali e ramificate verso il basso.
Date queste modalità di crescita, gli stadi 1-4 sono riconoscibili a prima vista per la presenza della freccia (assente negli stadi successivi) e per il profilo allungato, conico, della chioma.
Alla fine della fase giovanile la gemma apicale cessa l’attività di dominanza: la chioma sviluppatasi finora è definita temporanea (perché sarà in seguito eliminata per autopotatura), mentre quella definitiva inizierà a formarsi nello stadio 5.
Lo sviluppo dell’apparato radicale procede per stadi (indicati da lettere) parallelamente a quello della parte aerea (Fig. 1) ed è anch’esso programmato e regolato dalla dominanza esercitata dall’apice radicale.
Allo stadio A, il fittone inizia a crescere verticalmente e senza ramificazioni, inibite dalla dominanza apicale. Allo stadio B, si formano sottili radici laterali. Allo stadio C, dalla base del fittone si originano le principali radici fascicolate laterali. Allo stadio D, infine, il fittone si suddivide distalmente in diverse radici oblique principali, mentre alla base dello stesso si formano altre radici fascicolate, orizzontali od oblique. In sintesi, negli stadi A-D (corrispondenti agli stadi aerei 1-4) il compito principale è la costruzione del fittone (la radice centrale verticale).
Stadi 5 e 6: espansione in larghezza (Fig. 4)
In questa fase adulta la chioma si espande e l’albero acquisisce la sua dimensione massima. Il ritmo di crescita è decelerato, ma con una completa organizzazione della chioma in funzione del modello architettonico che caratterizza quella specie.
La freccia, ancora visibile all’inizio dello stadio 5, perde vigore e il suo apice non esercita più la dominanza sui rami sottostanti. Man mano che tale dominanza si indebolisce, l’apice delle branche situate nella parte alta della chioma, non più inibito, diviene dominante sulle ramificazioni della propria branca e, acquistato vigore, si raddrizza crescendo verso l’alto.
Tutta la periferia della parte alta della chioma acquista dunque vigore e forma la chioma definitiva: il profilo dell’albero, da conico, diventa arrotondato. Le branche più basse, sviluppatesi per parecchi anni sotto la forte dominanza apicale della freccia, tenderanno a seccare e a cadere (chioma temporanea: in grigio nella Fig. 4).
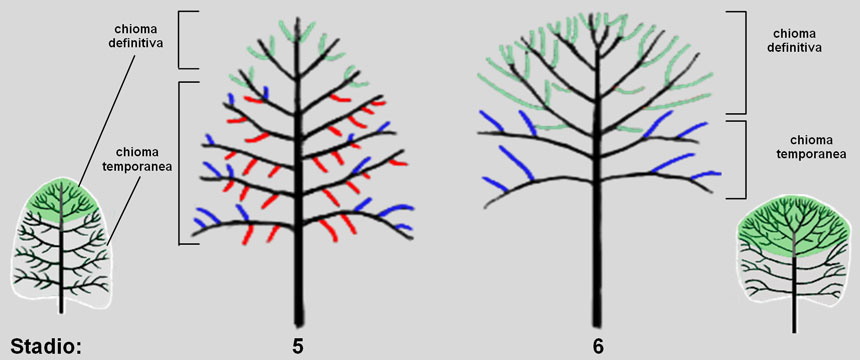 Fig. 4. Allo stadio 5 le nuove branche, non più sottoposte alla dominanza, possono svilupparsi verso l’alto e ramificarsi con modalità isotonica (cioè su tutti i lati: in verde nella figura), proprio come avveniva per le branche del tronco allo stadio 2. In pratica, ogni nuova branca si sviluppa come se fosse un nuovo tronco (reiterazione); da queste originerà la chioma definitiva, mentre le branche preesistenti –sviluppatesi sotto l’effetto della dominanza apicale– costituiscono la chioma temporanea (destinata a cadere). La chioma temporanea porta rami ipotoni (in rosso) e epitoni (in blu). Nello stadio 6 si completa la costruzione della chioma definitiva attraverso la successione di reiterazioni: ogni branca forma cioè un nuovo alberello figlio, le cui ramificazioni sviluppano nuovi alberelli e così via.
Fig. 4. Allo stadio 5 le nuove branche, non più sottoposte alla dominanza, possono svilupparsi verso l’alto e ramificarsi con modalità isotonica (cioè su tutti i lati: in verde nella figura), proprio come avveniva per le branche del tronco allo stadio 2. In pratica, ogni nuova branca si sviluppa come se fosse un nuovo tronco (reiterazione); da queste originerà la chioma definitiva, mentre le branche preesistenti –sviluppatesi sotto l’effetto della dominanza apicale– costituiscono la chioma temporanea (destinata a cadere). La chioma temporanea porta rami ipotoni (in rosso) e epitoni (in blu). Nello stadio 6 si completa la costruzione della chioma definitiva attraverso la successione di reiterazioni: ogni branca forma cioè un nuovo alberello figlio, le cui ramificazioni sviluppano nuovi alberelli e così via.
Allo stadio 5, dunque, ogni branca laterale superiore riproduce lo sviluppo già visto per il tronco: inizialmente la sua gemma apicale esercita la dominanza sulle gemme della branca stessa, permettendo lo sviluppo di ramificazioni secondarie deboli destinate a scomparire (ipotoni). In seguito, indebolendosi la dominanza, alcune gemme crescono verticalmente ramificandosi con modalità isotonica (rami figli disposti su tutti i lati del ramo genitore).
Queste nuove branche diventeranno i tronchi secondari costituenti l’ossatura della chioma definitiva e, come il tronco primario, subiranno un’intensa mortalità acropeta Acropeto: dal greco àcron ‘punta, cima, estremità’ e dal latino peto ‘tendere a’: che inizia cioè dalla base e procede verso l’estremità., si spoglieranno cioè dei propri rami basali.
In pratica, ogni branca sul tronco primario diventa indipendente dalla gerarchia precedentemente impostata e si sviluppa replicando la struttura e l’aspetto di un nuovo alberello (nuova unità architettonica), come se fosse un nuovo individuo: è il fenomeno della reiterazione. Gran parte del tronco porta ancora la chioma temporanea.
Nello stadio 6 si verifica una successione di reiterazioni totali: le grosse branche principali, infatti, si dividono in due o tre branche secondarie che crescono verso l’alto (sempre con modalità isotona) diventando a loro volta tronchi terziari di nuovi alberelli in cui la parte apicale del tronco manterrà la ramificazione che porta la chioma, mentre la parte basale si denuderà (per mortalità acropeta dei rami) (Fig. 5 e 6A).
Il ripetersi di reiterazioni sui nuovi rami che saranno generati fa sì che l’albero diventi una super-struttura gigante costituita da un albero “padre” che porta alberelli “figli” ciascuno dei quali porta “nipoti” ecc. Non si tratta solo di aspetto apparente, ma di vera sostanza anatomica e fisiologica, visto che il tronco di ogni “alberello” acquista gradualmente una certa indipendenza biologica dall’albero principale.
Ciascun “alberello”, infatti, è cioè dotato di propri fasci vascolari che, scendendo lungo i rami genitori, le branche e il tronco, raggiungono le proprie radici (sviluppatesi in continuità con essi). In diverse specie queste colonne legnose sono ben visibili al piede dell’albero, dove formano contrafforti (Fig. 6B e 6C), mentre in altre specie le colonne legnose non sono sporgenti, ma immerse nel tronco principale. Crescendo, dunque, l’albero perde la propria coesione e diventa un corpo composto da varie centinaia di individui.
L’insieme delle reiterazioni conduce all’espansione della chioma, al suo arrotondamento e al raggiungimento della dimensione massima dell’albero. Non si distingue più una linea del tronco, ma una serie di tronchi secondari che ramificano progressivamente (crescita simpodialeSimpodiale: tipo di ramificazione in cui l’asse principale ha accrescimento definito (destinato quindi ad arrestarsi) e produce lateralmente dei rami di primo, secondo, terzo ordine ecc., il cui accrescimento è via via maggiore nei rami successivi (pertanto il ramo di terz’ordine risulterà più lungo di quello di secondo ordine). Si contrappone a monopodiale.
Monopodiale: ramificazione di un albero che ha l’asse principale con crescita indefinita e rami di diversi ordini con sviluppo limitato e subordinati al tronco. In generale dicesi di un organo qualsiasi ad accrescimento indefinito. Si contrappone a simpodiale.). L’originaria ramificazione bassa, nata in una situazione di forte dominanza apicale (stadi 1-4), si sfronderà sempre più, mentre quella alta, nata in condizioni di dominanza apicale debole o assente, formerà la chioma definitiva.
 Fig. 5. Allo stadio 6 l’albero ha un’evidente organizzazione gerarchica, costituita da successive reiterazioni. In questo pioppo canadese, nonostante la copertura del fogliame, si riconoscono tre branche principali (lungo le linee tratteggiate 1-3) ognuna delle quali è una replica dell’albero, è cioè costituita da un tronco nudo e da varie ramificazioni che portano un’ampia chioma (segnalata dagli archi bianchi 1-3). A loro volta, dalle branche principali partono rami che sono anch’essi repliche in formato ridotto dell’albero: la loro chioma è segnalata dagli archi verde chiaro. Anche questi ultimi generano repliche in miniatura dell’albero, cioè rami nudi nella parte basale (equivalenti a un piccolo tronco) dai quali si dipartono ramoscelli frondosi (indicati dagli archi gialli).
Fig. 5. Allo stadio 6 l’albero ha un’evidente organizzazione gerarchica, costituita da successive reiterazioni. In questo pioppo canadese, nonostante la copertura del fogliame, si riconoscono tre branche principali (lungo le linee tratteggiate 1-3) ognuna delle quali è una replica dell’albero, è cioè costituita da un tronco nudo e da varie ramificazioni che portano un’ampia chioma (segnalata dagli archi bianchi 1-3). A loro volta, dalle branche principali partono rami che sono anch’essi repliche in formato ridotto dell’albero: la loro chioma è segnalata dagli archi verde chiaro. Anche questi ultimi generano repliche in miniatura dell’albero, cioè rami nudi nella parte basale (equivalenti a un piccolo tronco) dai quali si dipartono ramoscelli frondosi (indicati dagli archi gialli).
 Fig. 6. Esemplare di bagolaro (Celtis australis) in veste invernale lungo il viale XX Settembre, in località Gildona. Nell’albero spoglio sono meglio visibili le reiterazioni: senza il fogliame, infatti, è più facile vedere le singole ramificazioni. A: l’area delimitata dalla linea verde chiaro è una reiterazione (un alberello in miniatura) che, a sua volta, porta 4 reiterazioni più piccole (il cui fusto basale nudo, equivalente a un piccolo tronco, è indicato dalle frecce gialle). B: le specie dotate di contrafforti al piede (come il bagolaro) permettono di percepire meglio che questi sono la prosecuzione delle branche e dei rami principali lungo il tronco, che vanno a connettersi alle proprie radici. L’albero è dunque costituito dalla fusione di diversi alberelli in buona parte indipendenti, ciascuno dotato cioè di proprie radici e di un discreto grado di autonomia funzionale. C: come B, ma con l’indicazione dei singoli contrafforti in rilievo (linee verde chiaro) e della loro separazione (linee tratteggiate).
Fig. 6. Esemplare di bagolaro (Celtis australis) in veste invernale lungo il viale XX Settembre, in località Gildona. Nell’albero spoglio sono meglio visibili le reiterazioni: senza il fogliame, infatti, è più facile vedere le singole ramificazioni. A: l’area delimitata dalla linea verde chiaro è una reiterazione (un alberello in miniatura) che, a sua volta, porta 4 reiterazioni più piccole (il cui fusto basale nudo, equivalente a un piccolo tronco, è indicato dalle frecce gialle). B: le specie dotate di contrafforti al piede (come il bagolaro) permettono di percepire meglio che questi sono la prosecuzione delle branche e dei rami principali lungo il tronco, che vanno a connettersi alle proprie radici. L’albero è dunque costituito dalla fusione di diversi alberelli in buona parte indipendenti, ciascuno dotato cioè di proprie radici e di un discreto grado di autonomia funzionale. C: come B, ma con l’indicazione dei singoli contrafforti in rilievo (linee verde chiaro) e della loro separazione (linee tratteggiate).
Nello stadio 5 l’organizzazione della chioma è accompagnata dalla formazione completa dell’apparato radicale (stadio E, vedi Fig. 1): il fittone continua a svilupparsi in profondità e ad aumentare di diametro. Le radici fascicolate si sviluppano in larghezza, producendo sia sottili reiterazioni a rapida crescita, orizzontali od oblique, sia più vigorose reiterazioni verticali.
Nello stadio F (stadio aereo 6) l’apparato fittonante raggiunge il suo massimo sviluppo. Contemporaneamente, le reiterazioni fascicolate verticali si approfondiscono e vengono affiancate da nuove reiterazioni in posizione più periferica. Infine, alcune delle principali radici fascicolate orizzontali si orientano distalmente verso il basso formando “puntoni” di consolidamento.
Stadi 7 e 8: strategia di durata (Fig. 7)
La massima espansione della chioma è già stata acquisita nello stadio 6 e ormai le gemme apicali hanno cessato ogni attività di dominanza. Le ramificazioni della parte più esterna della chioma hanno ormai perso la capacità di crescere verticalmente (ortotropia) e non possono svilupparsi se non orizzontalmente (plagiotropia).
Nello stadio 7, pertanto, l’albero riduce il suo vigore e adotta una strategia di durata: per durare nel tempo è necessario rinnovare le ramificazioni man mano che deperiscono. Il principio generale seguito per il rinnovamento è la ramificazione per epitonia.
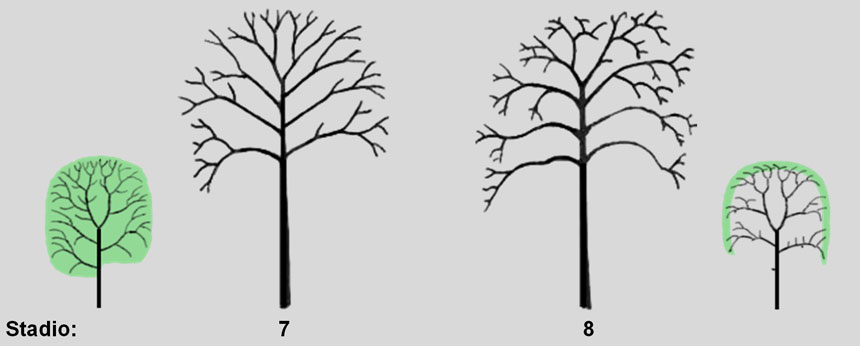 Fig. 7. Stadio 7: l’albero adotta una strategia di durata: la chioma non si espande ulteriormente, ma la sua ramificazione viene rinnovata per epitonia (si veda la Fig. 8). Stadio 8: la chioma perde vigore: è regolare e densamente ramificata alla sua periferia, ma si svuota all’interno.
Fig. 7. Stadio 7: l’albero adotta una strategia di durata: la chioma non si espande ulteriormente, ma la sua ramificazione viene rinnovata per epitonia (si veda la Fig. 8). Stadio 8: la chioma perde vigore: è regolare e densamente ramificata alla sua periferia, ma si svuota all’interno.
Man mano che un ramo si allunga e si incurva per il proprio peso, le sue ramificazioni cadono, iniziando da quelle basali e proseguendo verso l’estremità (mortalità acropeta). Quando il flusso di mortalità si avvicina all’estremità della ramificazione principale, quest’ultima viene sostituita da un ramo epitono: sul lato superiore germoglia cioè una gemma latente che sviluppa verso l’alto un ramo (più vigoroso del ramo principale) notevolmente ramificato; questo si accaparrerà gran parte dell’acqua e dei nutrienti provenienti dalle radici, provocando così l’indebolimento e la morte dell’estremità del ramo principale (Fig. 8).
In altre parole, il mantenimento della chioma avviene trasferendo la crescita dal ramo principale al suo ramo epitono, mentre i vecchi rami più distali sono progressivamente eliminati naturalmente (Fig. 8C). In seguito il ramo epitono, diventato ramo principale, origina una nuova ramificazione epitona che lo sostituisce e così via. In questo modo la chioma mantiene il suo volume complessivo, ma in un quadro di progressivo indebolimento: si rinnova continuamente alla periferia, ma si svuota al suo interno.
Il rinnovamento per epitonia è molto veloce e ben visibile nei tigli (Fig. 8E), mentre nella quercia è estremamente lento e si può apprezzare solo nell’arco di un secolo.
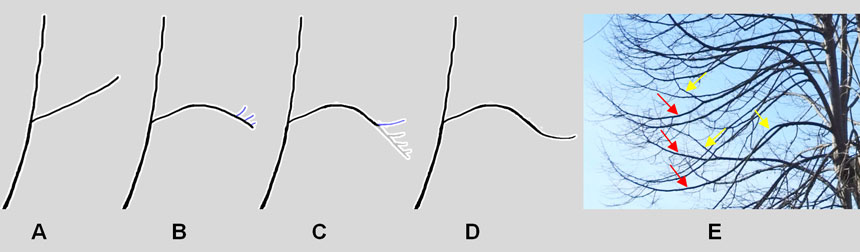 Fig. 8. Stadio 7: schema del rinnovamento dei rami per epitonia. Il ramo laterale (A), per l’effetto dell’allungamento e del suo stesso peso, tende a incurvarsi verso il basso e genera alcuni ramoscelli epitoni (rivolti verso l’alto: B). Uno di questi si sviluppa in un ramo vigoroso (ramo azzurro in C), mentre i rami più distali deperiscono progressivamente (rami grigi in C) e, in seguito, cadranno. D: il vigoroso ramo epitono ha sostituito quello caduto, rinnovando la chioma. E: nel tiglio ibrido in veste invernale sono ben individuabili i rami epitoni (frecce gialle) che sostituiranno la parte distale del ramo principale destinata a cadere (frecce rosse).
Fig. 8. Stadio 7: schema del rinnovamento dei rami per epitonia. Il ramo laterale (A), per l’effetto dell’allungamento e del suo stesso peso, tende a incurvarsi verso il basso e genera alcuni ramoscelli epitoni (rivolti verso l’alto: B). Uno di questi si sviluppa in un ramo vigoroso (ramo azzurro in C), mentre i rami più distali deperiscono progressivamente (rami grigi in C) e, in seguito, cadranno. D: il vigoroso ramo epitono ha sostituito quello caduto, rinnovando la chioma. E: nel tiglio ibrido in veste invernale sono ben individuabili i rami epitoni (frecce gialle) che sostituiranno la parte distale del ramo principale destinata a cadere (frecce rosse).
Allo stadio 8 si assiste all’inversione del flusso di mortalità, da acropeta a basipetaBasipeto: composto di basi- e –peto. Dicesi di organi o tessuti che si sviluppano alla base o che si dirigono verso la base dell’asse portante. È sinonimo di acrofugo e il contrario di acropeto e basifugo.
Acropeto: dal greco àcron ‘punta, cima, estremità’ e dal latino peto ‘tendere a’, che inizia cioè dalla base e procede verso l’estremità.: l’estremità dei rami va incontro a deperimento e viene sostituita da epitoni. Il processo si ripete negli anni, con sostituzione dell’estremità dei rami da parte di epitoni sempre più arretrati, il che porta ad una autoriduzione della chioma. Quest’ultima ha una forma molto omogenea, con un’alta densità di ramificazioni in periferia, mentre all’interno si spoglia (Fig. 7).
L’inversione del flusso di mortalità è accompagnata dunque dall’inversione della direzione di rinnovamento della chioma: i rami, infatti, si rinnovano (per epitonia) non più alla loro estremità ma, progressivamente, sempre più verso l’interno.
In questo stadio si registra dunque una notevole perdita di vigore della chioma e, alla fine, i nuovi getti annuali non riescono più a ramificarsi, ma crescono semplicemente al loro apice. Il motivo di questo indebolimento è stato attribuito al fatto che, col progressivo invecchiamento, l’albero aumenta la sua massa legnosa (per la formazione degli anelli annuali) cosicché, in proporzione, l’attività fotosintetica delle foglie diventa insufficiente all’accrescimento della massa legnosa e dell’apparato radicale, nonché ad accumulare le riserve necessarie a garantire una pronta risposta alle condizioni di stress.
Sembra tuttavia che la causa primaria stia nel deterioramento dell’apparato radicale, che non consentirebbe di sostenere un folto apparato fogliare. In effetti, nel passaggio tra gli stadi 7 e 8 si verifica una perdita massiva delle sottili radici assorbenti.
Lo stadio radicale G (stadio epigeo 7) è caratterizzato dal mantenimento delle potenzialità funzionali: le radici fascicolate raggiungono la loro massima espansione. Il fittone, pur ancora parzialmente vitale, risulta biologicamente ormai marginale.
A partire dallo stadio H (stadio aereo 8) inizia una consistente riduzione dell’apparato radicale: il fittone è ormai degradato e compare la mortalità delle radici fascicolate più profonde. L’apparato radicale diviene più superficiale e contratto, rendendo l’albero più sensibile agli stress ambientali.
Stadi 9 e 10: riduzione della chioma (Fig. 9)
Si è visto che allo stadio 8 il vigore dell’albero declina e il numero ridotto di foglie non è in grado di sostentare la considerevole quantità di legno accumulato fin dalla nascita. Nel cambioCambio: Tessuto meristematico, costituito da cellule giovani, posto tra lo xilema (legno) e il floema (libro, o cribro), nei fusti legnosi delle gimnosperme e nelle dicotiledoni.
Queste cellule, all’inizio della stagione vegetativa si moltiplicano e successivamente, differenziandosi, formano xilema e floema secondario.
In questo modo si formano gli anelli legnosi, dal cui conteggio è possibile stabilire l’età di un albero., inizialmente continuo, cominciano ad apparire dei vuoti; allo stesso tempo, in corrispondenza dei vigorosi rinnovi epitonici delle branche, la zona cambiale accresce la sua attività.
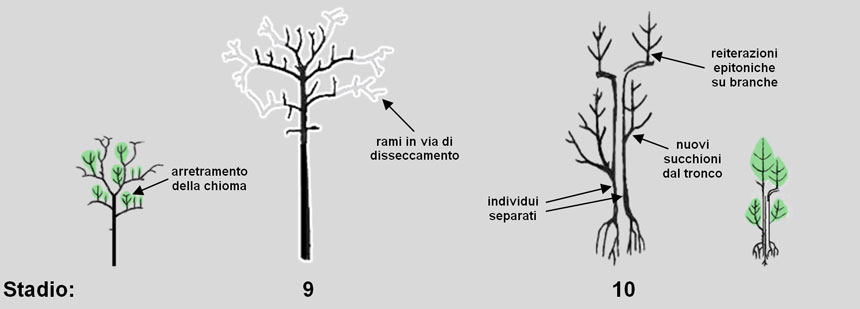 Fig. 9. Stadio 9: la chioma arretra progressivamente per morte (disseccamento) della parte distale dei rami (in grigio), sempre meno compensata da nuovi rami epitoni; al termine di questo stadio solitamente l’albero muore. Alcuni individui eccezionali raggiungono invece lo stadio 10 sviluppando vigorosi succhioni da gemme dormienti del tronco; il cambio genera una colonna legnosa e, raggiunto il suolo, un nuovo apparato radicale. Ne può risultare un tronco completamente cavo contornato da nuovi individui separati tra loro.
Fig. 9. Stadio 9: la chioma arretra progressivamente per morte (disseccamento) della parte distale dei rami (in grigio), sempre meno compensata da nuovi rami epitoni; al termine di questo stadio solitamente l’albero muore. Alcuni individui eccezionali raggiungono invece lo stadio 10 sviluppando vigorosi succhioni da gemme dormienti del tronco; il cambio genera una colonna legnosa e, raggiunto il suolo, un nuovo apparato radicale. Ne può risultare un tronco completamente cavo contornato da nuovi individui separati tra loro.
Nello stadio 9 prosegue l’abbassamento della cima per morte dei rami apicali, sostituiti da epitoni sempre più arretrati che svilupperanno una chioma più interna. Alla fine, questo arretramento conduce alla morte di grandi ramificazioni (solo parzialmente sostituite da nuove reiterazioni totali) e la chioma ne risulta ridotta, disorganizzata e con molti rami periferici secchi. In questo stadio si verifica la maggior percentuale di cedimenti dell’albero che, al termine dello stadio, solitamente muore.
In conseguenza della riduzione della chioma, l’apparato radicale (stadio J) decade velocemente perdendo prima gli elementi di ancoraggio verticale e, poi, riducendo la capacità esplorativa orizzontale per mortalità della porzione periferica dell’apparato fascicolato, parzialmente compensata dalla produzione di radici reiterate verticali collocate nella parte prossimale delle grandi radici fascicolate orizzontali.
Lo stadio 10 viene raggiunto solo da individui eccezionali. La chioma muore integralmente ma –in seguito alla caduta o al disseccamento di grosse branche– l’attività reiterativa, invece di cessare, può passare a ciò che resta del vecchio tronco primario, ormai del tutto cavo: da gemme latenti presenti nel tronco si sviluppano cioè alcune reiterazioni vigorose (succhioniI succhioni (volgarmente detti anche poppatoni) sono rami vigorosi diritti e a sviluppo verticale che nascono dai rami aerei o dal tronco (ma distanziati dal livello del suolo).
Poiché originano da gemme latenti (rimaste cioè in dormienza per lungo tempo), sorgono su rami e fusti già vecchi.
Sono chiamati così perché, a causa della loro conformazione, hanno una particolare capacità di risucchiare nutrienti dalla pianta, il che consente loro di svilupparsi in maniera molto veloce rispetto agli altri rami.
Poiché tendono ad assorbire molta linfa, rischiano di indebolire la pianta (peraltro, senza produrre né fiori, né frutti); in generale vanno perciò eliminati al momento della potatura. Differiscono dai polloni per il punto di origine.
I polloni, infatti, sono rami che nascono dalla base del tronco principale o dalle radici superficiali. Anch’essi sottraggono nutrimento alla pianta e vanno perciò eliminati, soprattutto nelle piante da frutto.).
Il cambio, divenuto ormai discontinuo, viene stimolato a intensificare la propria attività attorno alle ultime ramificazioni rimaste (i succhioni): qui, pertanto, il legno prodotto si sviluppa formando una colonna sporgente sul tronco che dà origine a un nuovo apparato radicale. Nello stadio 10 si verifica cioè una sorta di smembramento dell’albero in un sistema complesso di porzioni di albero che assumono una loro indipendenza, pur appoggiandosi in parte alla vecchia struttura portante.
Alla fine, pertanto, non si ha più un vero albero ma, lungo la circonferenza del tronco, diversi nuovi esemplari biologicamente indipendenti per l’approvvigionamento di acqua, minerali e carboidrati (Fig. 9 e 10). Negli alberi anziani si può avere una separazione fisica senza alcun danno per le parti vive dell’albero.
 Fig. 10. Il canforo ultrasecolare a fianco del municipio di Carrara è uno dei rari esemplari che ha raggiunto lo stadio 10. Il tronco è marcescente ma, in corrispondenza della vigorosa branca (freccia nera), il cambio ha formato una colonna legnosa (freccia blu) sporgente dal tronco e collegata a proprie radici sane (come rivela la chioma lussureggiante), tanto da risultare un individuo funzionalmente indipendente dal vecchio albero.
Fig. 10. Il canforo ultrasecolare a fianco del municipio di Carrara è uno dei rari esemplari che ha raggiunto lo stadio 10. Il tronco è marcescente ma, in corrispondenza della vigorosa branca (freccia nera), il cambio ha formato una colonna legnosa (freccia blu) sporgente dal tronco e collegata a proprie radici sane (come rivela la chioma lussureggiante), tanto da risultare un individuo funzionalmente indipendente dal vecchio albero.
Allo stadio K (stadio aereo 10), la generale riorganizzazione del cambio in corrispondenza dei nuovi succhioni sviluppatisi sul tronco stimola anche un progressivo incremento dell’attività reiterativa radicale. Le nuove radici si generano non solo alla base del vecchio tronco, ormai completamente cavo, ma anche in corrispondenza della corona. Guardando all’interno di un tronco cavo, infatti, sono spesso visibili molte radici aeree che appaiono sotto la base dei succhioni e scendono lungo il tronco fino ad affondarsi nel suolo.
L’apparato radicale è dunque ormai ridotto e funzionale solo alle porzioni vegetanti del relitto arboreo, che perde la sua unità tanto da non poter nemmeno essere considerato come un’unica struttura. In questa fase i cedimenti di rami diventano la normalità.
Lo sviluppo degli alberi: logica e implicazioni
Come si è visto, durante la crescita l’albero attraversa diversi stadi di sviluppo e muta aspetto seguendo uno schema generale, pur con differenze più o meno sensibili tra le singole specie. In queste righe cerchiamo di comprenderne la logica e le implicazioni.
Affinché l’albero possa svilupparsi nelle forme ordinate che conosciamo occorre un meccanismo gerarchico che stabilisca continuamente quali rami devono crescere, in quale direzione e per quale lunghezza e quali, invece, devono attendere il loro turno. In assenza di gerarchia, infatti, la ripetuta produzione di unità elementari (un internodo, un nodo e una gemma) in tutte le direzioni produrrebbe una massa sferica fittamente intricata (o, più esattamente, semisferica, visto che la crescita verso il basso è impedita dalla presenza del suolo).
Nella fase giovanile (stadi 1-4) il meccanismo gerarchico è la dominanza apicale (esercitata cioè dalla gemma apicale del tronco) che, attraverso la continua produzione di auxina, inibisce le gemme sottostanti costringendole a produrre rami in ritardo, con velocità ridotta e in direzione tendenzialmente orizzontale.
Anche le successive ramificazioni sono rivolte verso il basso o lateralmente. La crescita verso l’alto è ‘vietata’ ai rami per riservarla alla cima del tronco (la ‘freccia’) e consentire al giovane albero di crescere in altezza: un requisito fondamentale in natura, soprattutto nel bosco dove non vincere la corsa verso l’alto alla conquista della luce equivale spesso a una condanna a morte. Per questi motivi alla fine dello stadio 4 tutti gli alberi hanno la chioma conica e un profilo triangolare.
Nella fase adulta (stadi 5-6), terminata la dominanza esercitata dalla freccia, le nuove branche laterali possono finalmente crescere inclinate verso l’alto e ramificarsi (sempre verso l’alto) su tutti i lati. Le branche nate negli stadi precedenti (sotto l’effetto della dominanza) costituiscono la chioma provvisoria e vanno progressivamente incontro a disseccamento, morte e caduta, mentre le nuove branche producono ‘reiterazioni totali’, generano cioè nuovi ‘alberelli’ che, a loro volta, ramificandosi, ne generano altri formando una superstruttura che porta la chioma alle sue massime dimensioni in altezza e larghezza.
Ma questo processo non può proseguire all’infinito. La crescita in altezza è limitata dalla capacità dei vasi legnosi di trasportare acqua e nutrienti oltre una certa distanza, mentre la crescita in larghezza è limitata da problemi meccanici (un ramo troppo lungo e pesante tende a piegarsi e, poi, a spezzarsi).
Per questi motivi, una volta raggiunta la massima altezza e larghezza (negli stadi 5 e 6), nei successivi stadi 7 e 8 l’albero adotta una strategia di durata: la chioma viene rinnovata mediante la sostituzione dei rami che man mano deperiscono. Con gli anni, infatti, le branche primarie tendono progressivamente a piegarsi sotto il proprio peso: in tali condizioni, le loro ramificazioni rivolte verso l’alto si sviluppano e ramificano con maggior vigore di quelle sul lato inferiore e dell’estremità delle branche stesse. Le branche maggiormente sollecitate a piegarsi sono generalmente quelle più vecchie e lunghe, che si trovano nella parte bassa del tronco, e normalmente hanno le maggiori probabilità di rottura.
Si verifica così una selezione dei rami delle branche: quelli rivolti verso l’alto (epitoni), crescendo con maggior vigore, si accaparrano la linfa e progressivamente ‘affamano’ sia i rami ipotoni (rivolti in basso) sia l’estremità della branca stessa, che deperiscono e muoiono.
Si tratta del rinnovamento per epitonia: il vigoroso ramo epitono così selezionato sostituisce la parte distale della branca, destinata a cadere. Dopo anni, anch’esso sarà sostituito da un ramo epitono più interno, e così via per decenni. In questo modo i rami organizzano il rinnovo continuo di una parte sempre più importante della loro struttura.
Questo meccanismo è detto “post-selezione” (o selezione a posteriori) perché prima viene consentita la ramificazione e, solo dopo anni, vengono scelti i rami esistenti da eliminare perché si trovano in posizioni particolarmente sfavorite (ad es. troppo ravvicinati tra loro o all’interno della chioma o nella sua parte bassa, dove la mancanza di luce porta a un progressivo indebolimento e poi alla morte, oppure li costringe a un allungamento eccessivo fino a renderli meccanicamente fragili).
Perciò non tutti i rami nati da gemme in un dato anno potranno sopravvivere e continuare a produrre nuova vegetazione per sempre ma, a seconda dei casi, la pianta seguirà una strategia più o meno marcata di eliminazione e di semplificazione della propria struttura che porterà alla formazione di palchi (ad es. nel Cedro del Libano) o masse vegetative ben distinte che non si intralciano a vicenda nello sfruttamento della luce, oppure alla formazione di una chioma omogenea esternamente ma vuota all’interno, il tutto finalizzato ad ottenere la maggior superficie fotosintetica possibile con il numero minimo di ramificazioni.
Al contrario, nei primi stadi di sviluppo (1-4) si verifica la “pre-selezione” (o selezioni a priori): la selezione dei rami che non dovranno svilupparsi è infatti precocissima ed è ottenuta grazie alla dominanza apicale che inibisce la crescita delle loro gemme.
In alcuni alberi, come nella maggioranza delle conifere, lo stadio 4 ha una durata lunghissima: ciò spiega perché l’abete bianco e l’abete rosso, avendo una forte persistenza del germoglio apicale per gran parte della loro vita, mantengono la chioma conica per quasi tutta la vita. In questo caso, pertanto, la post-selezione è quasi assente in quanto la pianta adotta una rigida strategia di pre-selezione durante la formazione dei rami: la dominanza apicale costringe i meristemi a formare fin da subito una struttura gerarchicamente organizzata che in seguito non ha quasi bisogno di essere corretta.
La maggioranza degli alberi, invece, adotta la pre-selezione nella fase giovanile e, nell’età adulta, una forte selezione a posteriori, con la perdita progressiva delle ramificazioni più basse e la formazione di biforcazioni o cambi di direzione che dopo alcuni anni cambiano radicalmente l’aspetto della pianta.
Tale cambiamento di aspetto presenta indubbi vantaggi. Va infatti considerato che, nella loro vita plurisecolare, gli alberi devono riadattare il loro portamento all’orientamento della luce (magari perché la crescita di alberi vicini li ombreggia da un lato). La crescita indefinita e la post-selezione consentono pertanto di modificare nel tempo la loro chioma in modo da adattarla alle mutate condizioni ambientali: vento, temperatura, salsedine, smottamento del terreno, fulmini, incendi, concorrenza di altri alberi, brucatura di animali, attacchi di insetti, malattie, traumi ecc. La risposta a questi stress fa sì che ogni albero assume una forma unica e irripetibile.
Va infine segnalato che, pur seguendo lo schema generale descritto, alcune specie presentano peculiarità. Negli abeti, ad esempio, lo stadio 4 perdura per gran parte della vita. Il pino domestico, invece, è incapace al rinnovo della chioma per reiterazione epitoniche (tipiche dello stadio 7): ciò porta a un allungamento eccessivo delle branche che accentua la forma a ombrella della chioma, ma rappresenta anche una condanna meccanica (favorendone la caduta).
Aggiornamenti: